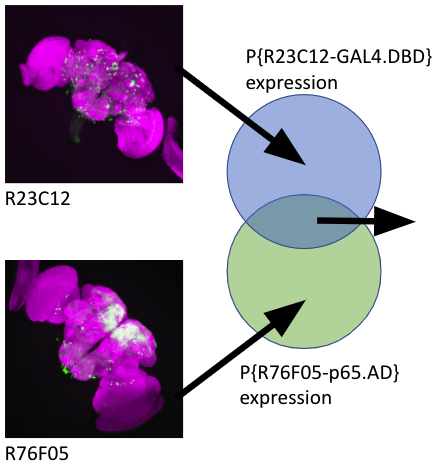
Insertion of 30kb into the coding region.
Insertion of about 30kb into the gl locus.
An insertion of about 30kb is within a 1.4kb XhoI fragment; position of restriction fragment on reference sequence inferred by FlyBase curator.
ommatidium | adult stage (with gl3)
photoreceptor cell & axon
photoreceptor neuron | P-stage (with gl3)
retina & axon
rhabdomere | adult stage (with gl3)
gl60j mutant flies fail to exhibit phototaxis from the day they eclose.
gl60j homozygous adults previously trained under 12h:12h light:dark cycles still maintain a rhythmic behavior under constant darkness conditions, but present a severe decrease in circadian phase shifting in response to light pulses during the late night period, as compared to controls.
Third instar eye discs in gl60j mutants display reduced numbers of photoreceptor neuron precursors and disorganized cellular arrangement, compared with controls. gl60j mutant photoreceptor precursors are not correctly recruited into the developing ommatidia in the third instar eye disc. Cellular organization of the retina in gl60j mutants remains severely compromised through metamorphosis to adulthood, although photoreceptor neurons remain present, in contrast to previous reports.
The cell bodies of photoreceptor neurons in gl60j mutants fail to elongate during metamorphosis.
The axonal projections of photoreceptor neurons in gl60j mutants are highly disorganized, branch profusely, and innervate primarily the medulla of the optic lobe. Histaminergic projections are still present but are disorganized as compared to wild-type. gl60j mutant photoreceptor neurons differentiate presynaptic specializations, but the synaptic vesicles are spread along the length of the axons, while in controls they accumulate at the distal tips of the axons. The lamina of the optic lobe is reduced in size in gl60j mutants, as compared to controls.
Large gl60j mutant clones in the developing eye do not acquire normal organization, fail to incorporate into the ommatidia, and re-localize to the basal side of the retina. This phenotype is not observed in small gl60j clones.
gl60j mutants, like wild type flies, show arrhythmic behavior in constant dim white light conditions.
gl60j/gl60j and gl3/gl60j mutants possess fewer and fused ommatidia, with no presence of differentiated rhabdomeres, and boundaries between ommatidia are difficult to distinguish, as compared to wild type.
Both gl60j/gl60j mutant larvae and gl60j/gl60j clones in the third instar eye imaginal disc display disrupted photoreceptor cluster patterning, but no aberrant or excessive cell death in the retina field, as compared to wild type. At 48 h APF, gl60j/gl60j mutants possess all expected cell types, photoreceptors, cone and pigment cells, as compared to wild type, however no ommatidia contains the correct number of each retinal cell type, there is complete disruption of organization of retinal cells into distinct identifiable ommatidia, and cone and primary pigment cells are either too few or too many in number, and there is some evidence of cell death in photoreceptor cells. At 72 h APF, there is extensive cell death in the retinal field and in photoreceptor cells.
At 96 h APF, gl3/gl60j mutants display increased cell death of photoreceptors in the retina.
gl60j mutants exhibit abnormal light-dependent temperature preference. In contrast to wild-type, they prefer higher temperature in the dark than in the light.
gl60j mutants not only have abnormal eye components, but also lack a subset of circadian clock cells, the posterior dorsal neurons (DN1[[p]]s).
gl60j mutants lack the majority of DN1 neurons except the two DN[1a]. The dorsal fusion commissure is thinner than in wild-type, probably as a result of loss of DNa projections.
When the locomotor activity of wild-type flies are monitored at 18 and 29oC, an earlier evening peak is seen at 18oC. This is not seen in mutants.
At high-light intensity, gl60j flies are able to resynchronize to a shift in the light-dark schedule of 8 hours. However, at 1000-fold lower light intensity, gl60j mutants take longer to synchronize to new schedules than wild-type flies. Entrained gl60j flies significantly anticipate the light-ON transient, while wild-type flies do not.
Unlike wild-type flies, gl60j flies do not show arrhythmia in response to constant light. These mutants are unable to entrain to red light-dark schedules, to which wild-type flies can entrain.
gl60j mutants lack the ocelli, the ommatidial photoreceptors, the primary and secondary pigments of the eye, Bolwigs organ and the eyelet.
Third instar larvae have detectable dendritic arborisation of the lateral neurons in only 18% of cases and the average area of the arborisation is reduced compared to controls.
Homozygous flies lack the Hofbauer-Buchner (H-B) eyelet cells. The morphology of the optic lobes is altered. There is a reduction in the Pdf-positive arborisations in the accessory medulla and the medulla. Mutant flies can entrain and reentrain to 12 hour light-12 hour dark (LD) cycles. The morning peak is less compact than in wild-type flies, is not as well entrained to the LD cycles and takes an average of 6 days to adapt to the new phase, whereas the evening peak adapts quickly. Flies show 100% rhythmicity under LD conditions. Most flies become arrhythmic under constant light conditions.
Wild-type larvae reduce their path lengths when exposed to light. This response is severely reduced in gl60j larvae. Wild-type larvae show increased head swinging when exposed to light. This response is abolished in gl60j larvae. Wild-type larvae show a greater change in direction when lights are turned on or off (light (L) to dark (D), or D to L transition) than in the absence of a light transition (D to D). The amplitude of change of direction is greater for the D to L than for the L to D transition. This difference in the amplitude of change of direction between the D to L and L to D transitions is abolished in gl60j larvae, although change of direction in the absence of a light transition (D to D) is still significantly lower than for either the D to L and L to D transition.
Only a few loosely clustered photoreceptors can be seen without formation of a larval optic nerve. LP1 cells show abnormal neuronal processes but no clear Ddc arborisation can be detected.
Eye phenotype.
Third instar foraging and wandering stage larvae fail to respond to light.
Displays locomotor activity rhythm with an approximately 24h period.
Pattern of retinal axon projections in the larval brain is completely disorganised. Axon fascicles take aberrant trajectories after they exit the optic stalk and terminate in ectopic locations often outside the lamina target area.
Photoreceptor axons project to the brain aberrantly in late third instar larvae. This phenotype is variable. There is a variable pattern of S phase cells located between the outer proliferative centre and inner proliferative centre, in the region where the lamina precursor cells normally reside.
Strong allele. The eye is disorganised. Corneal hairs and red pigment granules are only present in the centre of the eye. The ocellar lenses are normal in size, but lack the bulge of the wild-type, and have small protuberances. The lenses of the compound eye and ocelli are roughened with "corneal nipples".
gl60j has photoperiod response variant | adult stage phenotype, enhanceable by Rh71/Rh71
gl60j has abnormal locomotor rhythm phenotype, enhanceable by cryb
gl60j has increased cell death | P-stage phenotype, suppressible by BacA\p35UAS.cUa/Scer\GAL4so.PU
gl60j has increased cell death | P-stage phenotype, suppressible by BacA\p35UAS.cUa/Scer\GAL4hs.PU
gl60j has visible | adult stage phenotype, non-suppressible by Pph13hs.PL
gl60j has visible | adult stage phenotype, non-suppressible by BacA\p35UAS.cUa/Scer\GAL4so.PU
gl60j has visible | adult stage phenotype, non-suppressible by BacA\p35UAS.cUa/Scer\GAL4hs.PU
gl60j/gl60j is an enhancer of photoperiod response variant | adult stage phenotype of Rh71
gl60j is an enhancer of abnormal circadian rhythm phenotype of so1
gl60j is a suppressor of photoperiod response variant phenotype of cryb
gl60j/gl[+] is a suppressor of visible phenotype of Pp2B-14Dact.GMR
gl60j is a non-suppressor of photoperiod response variant phenotype of Pdf01
gl60j is a non-suppressor of abnormal locomotor rhythm phenotype of Pdf01
Rh71, gl60j has abnormal circadian rhythm | adult stage phenotype
cryb, gl60j has abnormal locomotor rhythm phenotype
gl60j has eye photoreceptor cell | P-stage phenotype, suppressible by glhs.PL/Pph13hs.PL
gl60j has rhabdomere phenotype, suppressible by glhs.PL/Pph13hs.PL
gl60j has ommatidium | adult stage phenotype, non-suppressible by BacA\p35UAS.cUa/Scer\GAL4hs.PU
gl60j has eye photoreceptor cell | adult stage phenotype, non-suppressible by BacA\p35UAS.cUa/Scer\GAL4so.PU
gl60j has eye photoreceptor cell | adult stage phenotype, non-suppressible by BacA\p35UAS.cUa/Scer\GAL4hs.PU
gl60j has eye photoreceptor cell | P-stage phenotype, non-suppressible by Pph13hs.PL
gl60j has eye phenotype, non-suppressible by Pph13hs.PL
gl60j has eye phenotype, non-suppressible by BacA\p35UAS.cUa/Scer\GAL4so.PU
gl60j has eye phenotype, non-suppressible by BacA\p35UAS.cUa/Scer\GAL4hs.PU
gl60j has ommatidium | adult stage phenotype, non-suppressible by BacA\p35UAS.cUa/Scer\GAL4so.PU
gl60j/gl[+] is a suppressor of eye phenotype of Pp2B-14Dact.GMR
Rh71, gl60j double homozygous adults previously trained under 12h:12h light:dark cycles display both an increased proportion of arrhythmic individuals under constant darkness conditions and increased delay in circadian phase shifting to a 8h delay in the light:dark transition, as compared to either single homozygote.
Pulsing Pph13hs.PL expression using heat shocks does not rescue photoreceptor differentiation in gl60j/gl60j mutant retinas at 96 h APF, and does not further disrupt or rescue overall eye morphology of gl60j/gl60j mutants.
Pulsing expression of both Pph13hs.PL and glhs.PL using heat shocks restores photoreceptor differentiation in gl60j/gl60j mutant) retinas, as indicated by the presence of definable rhabdomeres at 96 h APF.
gl60j; so1 double mutants are completely eyeless and the eyelet is absent. These double mutants can only re-entrain to wavelengths shorter than 450nm, while so1 single mutants can entrain to wavelengths up to 550nm. Unlike so1 single mutants, phase relationship the evening peak activity of gl60j; so1 double mutants is not dependent on wavelength and these flies rarely show antidromic phase shifting.
The rough eye phenotype of Pp2B-14Dact.GMR flies is strongly suppressed by one copy of gl60j.
gl60j cryb double mutant flies do not entrain at all to 12 hour light-12 hour dark (LD) cycles. At 100 lux, approximately 70% of the flies show free-running behaviour. The average activity shows a small peak after lights off at 100 lux and at 1000 lux virtually all flies react to lights off with a pronounced activity peak. At 1000 lux, the activity level is generally lower during the light phase than during the dark phase. A free-running circadian activity component is present in 81% of the activity records, and after transfer into constant darkness conditions rhythmic locomotor behaviour usually continues from this free-running component and not from the lights-off peak. Approximately 30% of gl60j cryb double mutant flies show poor rhythmicity, even under LD conditions. At 100 lux, about 20% of the flies are arrhythmic, and about 10% show a complex rhythmicity characterised by several small but significant peaks in the periodograms. gl60j cryb double mutants show neither period changes nor any increase in activity level or intervals between morning and evening locomotor activity peaks after transfer from constant darkness to constant light conditions.
Weak suppressor of the rough eye phenotype caused by two insertions of P{GMR-Rho1}.
Expression of BacA\p35Scer\UAS.cUa under the control of Scer\GAL4so.PU or Scer\GAL4hs.PU results in a decrease in cell death at 48 h APF, but did not rescue the eye morphology, retina structure, or photoreceptor differentiation defects of gl60j/gl60j mutants.
Periodic pulses after 24 h APF of glhs.PL using heat shocks neither rescues or induces additional defects with respect to eye morphology, but restores photoreceptor differentiation in gl60j/gl60j mutant retinas, as indicated by the presence of rhabdomeres separated by a distinct inter-rhabdomeral space, although many of these ommatidia still do not contain the full complement of eight photoreceptors.
Expression of a single copy of glT:Avic\GFP-SF,T:Zzzz\FLAG,T:Zzzz\PSP.CS,T:Zzzz\TEV.CS,T:Zzzz\BLRP rescues the overall external morphology of the eye, but not the photoreceptor differentiation defects of adult gl3/gl60j and gl60j/gl60j mutants. In gl60j/gl60j mutants at 48 h APF, expression of a single copy of glT:Avic\GFP-SF,T:Zzzz\FLAG,T:Zzzz\PSP.CS,T:Zzzz\TEV.CS,T:Zzzz\BLRP significantly increases the number of ommatidia with the correct numbers of photoreceptor and cone cells.
Expression of two copies of glT:Avic\GFP-SF,T:Zzzz\FLAG,T:Zzzz\PSP.CS,T:Zzzz\TEV.CS,T:Zzzz\BLRP rescues the external morphology of the eye and photoreceptor differentiation defects of gl60j/gl60j mutants.